 Evolution
Evolution
Fact-Checking Wikipedia on Common Descent: The Evidence from Paleontology
Previously in this series, I have discussed the arguments for common descent presented by Wikipedia in the areas of comparative physiology/biochemistry and comparative anatomy. In this third installment, I will discuss the arguments from paleontology and the fossil record. Wikipedia’s page on the “Evidence of common descent” contains only a brief section on the fossil record, half of which is concerned with describing the discipline of paleontology and noting the limitations of the fossil record.
Wikipedia correctly observes that “because of limitations inherent in the record, there are not fine scales of intermediate forms between related groups of species. This lack of continuous fossils in the record is a major limitation in tracing the descent of biological groups.” The page subsequently lists nine rationalizations for the alleged incompleteness of the fossil record. It should be noted, however, that offering a rationalization for why the fossil record does not demonstrate a continuous progression of forms is not the same as using the fossil record as evidence for that same phenomenon.
Since this section of Wikipedia’s page does not really discuss the fossil evidence for common descent (except for the evolution of the horse), I will comment primarily on the page to which Wikipedia links, entitled “Transitional fossil“.
Exhibit A: Archaeopteryx
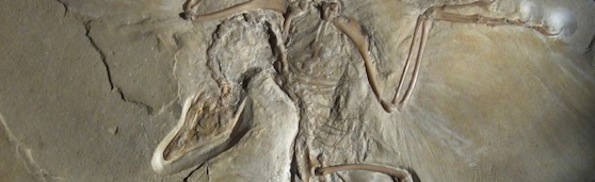
The first “transitional fossil” that Wikipedia mentions is Archaeopteryx, and we are informed that “Archaeopteryx was discovered just two years later [than the publication of The Origin], in 1861, and represents a classic transitional form between dinosaurs and birds.” Features that once convinced paleontologists of Archaeopteryx‘s transitional status with respect to theropod dinosaurs and birds include the fact that it possessed fully developed feathers (including primaries and secondaries) which were arranged on its wings in a manner similar to modern birds, as well as its reptilian features such as its long bony tail, the claws on its forelimbs and hindlimbs, and its teeth. Other fossilized birds, however, have been found to have teeth (e.g. Hesperornis) and, indeed, some modern birds (e.g. the ostrich) possess fingers on their wings.
Archaeopteryx cannot have descended from the so-called feathered theropod dinosaurs (which exhibit the most bird-like characteristics) because they lived some tens of million years after Archaeopteryx. Actually, these feathered dinosaurs emerge even later than various groups of Cretaceous birds which were more similar to modern ones than was Archaeopteryx. Anyone who has carefully followed the scientific discussion on evolution and the fossil record will doubtless be aware that Archaeopteryx has since been knocked off its perch, as it were, as the first bird, following the discovery of Xiaotingia zhengi in western Liaoning, China, in rocks dating to the late Jurassic epoch, 161-145 million years ago (Xu et al., 2011). As one news report in Nature put it, “we are about to enter a new era in which Archaeopteryx is considered as distant from the ancestry of modern birds as dinosaurs such as Deinonychus.” This is, of course, not the first time that the bird-like status of Archaeopteryx has been called into question. Indeed, one 2009 report stated:
The feathered creature called archaeopteryx, easily the world’s most famous fossil remains, had been considered the first bird since Charles Darwin’s day. When researchers put its celebrity bones under the microscope recently, though, they discovered that this icon of evolution might not have been a bird at all.
An examination of its bone cells revealed for the first time that the 150-million-year-old creature had the slow growth rate of a dinosaur, not a bird, an international research team reported this month. Comparing it with other early fossils, the researchers concluded that the telltale physiology of modern birds likely didn’t emerge until 20 million years or so after archaeopteryx flapped its broad wings across primordial lagoons.
As another news story on the new find admitted, however,
The researchers acknowledged that their reclassification was “only weakly supported by the available data,” but they said this kind of fuzziness was to be expected when the fossils being analyzed are close to the common ancestor of now-extinct dinosaurs and modern birds. “This phenomenon is also seen in some other major transitions, including the origins of major mammalian groups,” they wrote.
Witmer agreed: “We’re looking at an origin, and consequently it’s going to be messy.” […] “It just shows what evolution is all about. A prediction of evolutionary theory is that it should be really hard for us to figure out what’s going on in an origin.”
Indeed, the even earlier date for the emergence of birds only further accentuates the problem of their having preceded the nonflying feathered theropod dinosaur fossils, which have traditionally been seen as the ancestors of birds. Moreover, one consideration that is often overlooked in these discussions is the numerous problems with explicating a mechanism that could facilitate such a transition. Such problems include the following:
- In order to support the very high metabolism associated with flying, birds have a unique manner in which they distribute air around their bodies. There is zero evidence, however, of reptiles ever having such a system or anything that may be remotely considered its precursor. Indeed, one study published in Science concluded that the theropods had a crocodilian mode of breathing, and thus to convert this to the bird system would require passing through a non-viable transitional stage (Ruben et al., 1997).
- The theropods were Saurischian, and not Ornithiscian: That is, they had a distinctive lizard-like pelvis, rather than a bird-like pelvis.
- Both birds and theropod dinosaurs possess three digits in their forelimbs, but they are different ones (second to fourth and first to third respectively).
- The forelimbs of birds must be able to support their wings and are thus well developed. But the theropods possessed very small forelimbs, which are hardly credible precursors to those of birds and, indeed, a creature like Archaeopteryx.
- Presumably indicative of the requirement for increased sensory input and motor control for flying, Archaeopteryx possessed a larger braincase.
Indeed, as ENV’s Casey Luskin notes here, scientists now increasingly question the viability of the theropod-to-bird theory. As one 2009 article on Science Daily reported,
It’s been known for decades that the femur, or thigh bone in birds is largely fixed and makes birds into “knee runners,” unlike virtually all other land animals, the [Oregon State University] experts say. What was just discovered, however, is that it’s this fixed position of bird bones and musculature that keeps their air-sac lung from collapsing when the bird inhales.
Warm-blooded birds need about 20 times more oxygen than cold-blooded reptiles, and have evolved a unique lung structure that allows for a high rate of gas exchange and high activity level. Their unusual thigh complex is what helps support the lung and prevent its collapse.
“This is fundamental to bird physiology,” said Devon Quick, an OSU instructor of zoology who completed this work as part of her doctoral studies. “It’s really strange that no one realized this before. The position of the thigh bone and muscles in birds is critical to their lung function, which in turn is what gives them enough lung capacity for flight.”However, every other animal that has walked on land, the scientists said, has a moveable thigh bone that is involved in their motion — including humans, elephants, dogs, lizards and — in the ancient past — dinosaurs.
The implication, the researchers said, is that birds almost certainly did not descend from theropod dinosaurs, such as tyrannosaurus or allosaurus. The findings add to a growing body of evidence in the past two decades that challenge some of the most widely held beliefs about animal evolution.“For one thing, birds are found earlier in the fossil record than the dinosaurs they are supposed to have descended from,” Ruben said. “That’s a pretty serious problem, and there are other inconsistencies with the bird-from-dinosaur theories.
“But one of the primary reasons many scientists kept pointing to birds as having descended from dinosaurs was similarities in their lungs,” Ruben said. “However, theropod dinosaurs had a moving femur and therefore could not have had a lung that worked like that in birds. Their abdominal air sac, if they had one, would have collapsed. That undercuts a critical piece of supporting evidence for the dinosaur-bird link.
A further difficulty pertains to explaining the origin of feathers. If, as is conventionally maintained, feathers evolved from scales, one has to posit some kind of Darwinian explanation for their evolution from perhaps frayed scales, which must include some kind of genetic basis for every step from frayed scale to fully sculptured feather complete with its many specialized features. Based on what we presently know of the genetic mechanisms, structure and molecular composition of functioning feathers, the proposition that these structures evolved from frayed scales may now reasonably be regarded as ludicrous.
Exhibit B: Horse Evolution
The next specimen to be mentioned by Wikipedia is the horse series. The subject is covered in more detail on the “Evidence of common descent” page. We are told,
Due to an almost-complete fossil record found in North American sedimentary deposits from the early Eocene to the present, the horse provides one of the best examples of evolutionary history (phylogeny).
This evolutionary sequence starts with a small animal called Hyracotherium (commonly referred to as Eohippus) which lived in North America about 54 million years ago, then spread across to Europe and Asia. Fossil remains of Hyracotherium show it to have differed from the modern horse in three important respects: it was a small animal (the size of a fox), lightly built and adapted for running; the limbs were short and slender, and the feet elongated so that the digits were almost vertical, with four digits in the forelimbs and three digits in the hindlimbs; and the incisors were small, the molars having low crowns with rounded cusps covered in enamel.The probable course of development of horses from Hyracotherium to Equus (the modern horse) involved at least 12 genera and several hundred species. The major trends seen in the development of the horse to changing environmental conditions may be summarized as follows:
- Increase in size (from 0.4 m to 1.5 m — from 15in to 60in);
- Lengthening of limbs and feet;
- Reduction of lateral digits;
- Increase in length and thickness of the third digit;
- Increase in width of incisors;
- Replacement of premolars by molars; and
- Increases in tooth length, crown height of molars.
Fossilized plants found in different strata show that the marshy, wooded country in which Hyracotherium lived became gradually drier. Survival now depended on the head being in an elevated position for gaining a good view of the surrounding countryside, and on a high turn of speed for escape from predators, hence the increase in size and the replacement of the splayed-out foot by the hoofed foot. The drier, harder ground would make the original splayed-out foot unnecessary for support. The changes in the teeth can be explained by assuming that the diet changed from soft vegetation to grass. A dominant genus from each geological period has been selected to show the slow alteration of the horse lineage from its ancestral to its modern form.
Actually, the ancestry of the horse is one of the best indications we have that diversification/specialization is based predominantly upon the segregation of genes already present in a primordial group, rather than upon the acquisition of genes. Based on the fossil record, the horse series looks something like this:
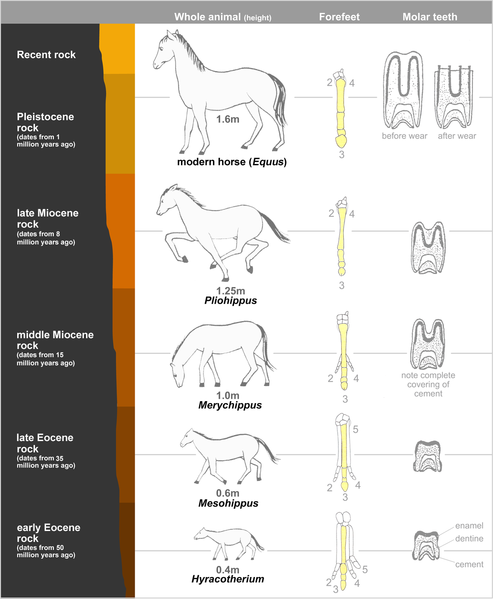
Hyracotherium (sometimes known as Eohippus) was fully herbivorous: its intestines included a caecum which accommodated bacteria for digesting cellulose and other plant material. Its teeth also corroborate an herbivorous diet. It was lightly built, with a tip-toed gait and slender legs (making it efficient as a runner). All subsequent modifications of Hyracotherium involved fairly minor tweaking of the characters present in this animal, comparable with the diversity among different breeds of dog. An indication that these differences were quite minor is the fact that all of these species are classified within one family (Equidea). Moreover, while the common vision is of horses becoming gradually larger over time with a trend toward increasing footbone length, this is true only of some lines, whereas others become smaller and, in some lineages, the footbones also become smaller.
Features that are used to discriminate among the different genera include the single-toed ligament, various features associated with the teeth (e.g. high crowns), variations of the skull (e.g. the disappearance of the preorbital fossa and the retraction/deepening of the nasal opening). Particularly remarkable, however, is that these features appear to have recurred multiple times in independent horse lineages, even in rather different environments (e.g. see Forsten, 1989). For example, both the European line and the North American line developed some of the same characteristics.
The two most plausible candidate explanations for this, in my opinion, are:
1. The necessary genes were already present in the original Hyracotherium population.
2. The diversification may have arisen by virtue of gene segregation.
In other words, from the beginning to the end of the series, we’re looking at relatively small-scale microevolutionary change. Let’s examine in turn the evidence for these two explanations.
In the case of the former, one possible piece of supportive evidence is that various species of Hipparion, though still possessing three “primitive” toes, nonetheless also possessed the ligament system associated with single toes. Given that the single-toe ligament system is meant to have been an adaptation to possession of a single toe, it seems somewhat unlikely that it would have evolved in three-toed animals.
As for the second possible explanation, all of the early European lineages of horses become extinct by the close of the Oligocene. This may be a result of the fact that the original genetic diversity had been segregated and thus the individual horse genera possessed limited genetic adaptability and variability, making them more susceptible to extinction. In contrast, the horses in North America, which did not change much during the Eocene or Oligocene, retained their variability and did not face such extinction. Having subsequently diversified from the beginning of the Miocene, however, most of these North American lineages faced extinction by the close of the Miocene — perhaps suggesting a common cause.
Exhibit C: Whale Evolution

The next transitional series alluded to by Wikipedia is the whale series. One of the most notable problems with the evolution of the whale is the extremely abrupt timescale over which it is supposed to have occurred. The sheer force of this conundrum is only properly appreciated when one considers the multiple feats of anatomical novelty, innovative engineering and genetic rewiring necessary to change a terrestrial mammal like Pakicetus into a fully aquatic whale. Indeed, evolutionary biologist Richard Sternberg has argued that even many of the relatively minor changes are extremely unlikely to have occurred in the time-frame allowed. Consider the following small sample of necessary modifications:
- Counter-current heat exchanger for intra-abdominal testes
- Ball vertebra
- Tail flukes and musculature
- Blubber for temperature insulation
- Ability to drink sea water (reorganization of kidney tissues)
- Fetus in breech position (for labor underwater)
- Nurse young underwater (modified mammae)
- Forelimbs transformed into flippers
- Reduction of hindlimbs
- Reduction/loss of pelvis and sacral vertebrae
- Reorganization of the musculature for the reproductive organs
- Hydrodynamic properties of the skin
- Special lung surfactants
- Novel muscle systems for the blowhole
- Modification of the teeth
- Modification of the eye for underwater vision
- Emergence and expansion of the mandibular fat pad with complex lipid distribution
- Reorganization of skull bones and musculature
- Modification of the ear bones
- Decoupling of esophagus and trachea
- Synthesis and metabolism of isovaleric acid (toxic to terrestrial mammals)
- Emergence of blowhole musculature and their neurological control
According to Richard Sternberg’s calculations, and based on the equations of population genetics applied in a 2008 paper by Durrett and Schmidt in the Journal of Genetics, one may reasonably expect to see two co-ordinated mutations to achieve fixation in the timeframe of around 43.3 million years. When one considers the magnitude of the engineering feat, such a scenario can only be ruled incredible. This problem is accentuated further when one considers that the majority of anatomical novelties unique to aquatic cetaceans (Pelagiceti) appeared during just a few million years — probably within 1-3 million years.
Until recently, the whale series, going from Pakicetus (53 mya) to Dorudon and Basilosaurus (38 mya) has looked something like this:
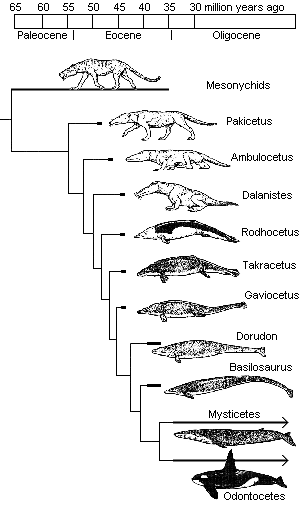
More recently, however, a jawbone was discovered that belonged to a fully aquatic whale dating to 49 million years ago, only four million years after Pakicetus! This means that the first fully aquatic whales now date to around the time when walking whales (Ambulocetus) first appear. This substantially reduces the time window — to 4 or 5 million years, perhaps even less — that may be allotted to the Darwinian mechanism to accomplish truly radical engineering innovations and genetic rewiring. It also suggests that this fully aquatic whale existed before its previously presumed semi-aquatic archaeocetid ancestors.
Exhibit D: Tiktaalik

The next example given of a transitional fossil is the infamous Tiktaalik fossil, the supposed intermediate between fish and amphibian tetrapods. This interpretation, however, has faced substantial problems since a study published in Nature last year reported on the discovery of tetrapod footprints dated to 395 million years ago, substantially preceding Tiktaalik (which dates to some 375 million years ago). See this popular-level Nature news report for more.
Furthermore, as ENV’s Casey Luskin discusses here, evidence of a transition connecting the ray-finned fish-fin of Tiktaalik and the true tetrapod limbs of Acanthostega and Tulerpeton is sorely lacking.
A Curious Omission: The Cambrian Explosion
Wikipedia curiously omits any discussion of the numerous problems for evolutionary theory that arise from the Cambrian explosion, a monumental event in the history of life where the majority of the major animal groups (the phyla) first appear — very abruptly — in the fossil record, with no observable intermediates. One of the most frequent ways of attempting to circumvent this challenge is to posit that organisms prior to the Cambrian were of a non-fossilizable composition. And, indeed, it is to be expected that non-skeletonized predecessors would leave few (if any) fossils. If it were the case, therefore, that one evolving line appeared suddenly in the fossil record, once it reached the stage of being fossilizable, then the point would have some validity. But the real challenge posed by the Cambrian explosion comes from the wide variety of fossilizable forms that appeared at more or less the same instant in geological time. Most of the major phyla represented by modern day organisms — certainly all those with fossilizable parts — were included, yet for none is there any clearly identifiable ancestor. Explaining this simultaneous and abrupt appearance is one of the leading challenges in evolutionary biology.
Interestingly, over the last century and a half since the publication of Darwin’s Origin of Species, paleontologists have discovered many Precambrian fossils, many of them microscopic or soft-bodied. As Darwinian paleobiologist William Schopf wrote in his The early evolution of life: solution to Darwin’s dilemma, “The long-held notion that Precambrian organisms must have been too small or too delicate to have been preserved in geological materials…[is] now recognized as incorrect.” If anything, the abrupt appearance of the major animal phyla, conventionally dated to about 540 million years ago, is better documented now that in Darwin’s time. Indeed, with an enhanced fossil record comes the realization that the Cambrian radiation was more explosive and abrupt than had been previously thought.
At any rate, as discussed in some detail here, the Ediacaran fauna are not broadly viewed as being the ancestors of the modern taxonomic groups (such as the various phyla which appear abruptly in the Cambrian radiation). The presence of these organisms, therefore, should offer no comfort to Darwinists. As Peter Ward has observed in On Methuselah’s Trail: Living Fossils and the Great Extinctions,
[L]ater study cast doubt on the affinity between these ancient remains preserved in sandstones and living creatures of today; the great German paleontologist A. Seilacher, of T�bingen University, has even gone so far as to suggest that the Ediacaran fauna has no relationship whatsoever with any currently living creatures. In this view, the Ediacaran fauna was completely annihilated before the start of the Cambrian fauna.
Moreover, many phyla (such as the brachiopods and arthropods) couldn’t have evolved their soft parts first and then added the hard parts (such as the exoskeleton or shell) later — their survival depends in large measure upon the ability to protect or shield their soft parts. Soft and hard parts had to arise together.
Summary & Conclusion
It is a popular view within evolutionary biology that the fossil record provides key evidence for common ancestry. But no phylum can be traced from a previous phylum in the fossil record. Indeed, this phenomenon holds with regard to the lower taxonomic categories (e.g. classes and orders). While transitional series can, in some cases, be found with respect to genera and species, the scope for change is clearly limited. And in instances that are often thought to be the best-case scenarios in the fossil record (e.g., the evolution of the whale), the transition occurs far too abruptly to be reasonably attributed to a blind Darwinian process. The fossil record, far from establishing Darwinian theory, only succeeds in hammering one more nail into Darwin’s coffin. In the next installment of this series, I will discuss the arguments for common descent from biogeographical distribution.