 Evolution
Evolution
 Intelligent Design
Intelligent Design
Laurence Moran’s Sandwalk Evolves Chloroquine Resistance
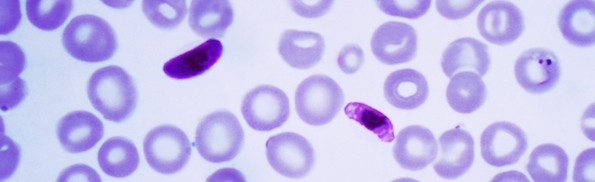
First, a bit of background. As I discussed previously (see here, here, and here) in a new paper Summers et al. show that a minimum of two mutations to the malarial protein PfCRT are needed to confer an ability to pump the antibiotic chloroquine (which is necessary but may not be sufficient for chloroquine resistance in the wild).
That result agrees with my discussion in The Edge of Evolution and goes a long way toward quantitatively explaining the rarity of the development of resistance. Over at PZ Myers’s blog Pharyngula, he and Kenneth Miller disagreed strongly with me in words, but cited no numbers. I then invited them, since they don’t like mine, to show us their calculations for how frequently chloroquine resistance should arise in the malarial parasite.
The bad news is that so far neither has responded. The good news is that Laurence Moran, Professor of Biochemistry at the University of Toronto, has done so. Professor Moran is an intelligent, informed, direct, and relatively civil critic of intelligent design who maintains a popular blog, Sandwalk, on evolution-related matters. So his response gives us a great opportunity to see what the best alternative explanations might be.
Moran begins his own calculation by assuming that the first required mutation is strictly neutral and spreads in the growing population before the second one arises. His straightforward computation leads him to conclude that "What this means is that if you start with an infection by a cell that has none of the required mutations then you will only get the right pair of mutations once in one million infected people."
Once in one million infected people…. Since there are a trillion malarial cells in one sick person, then according to Moran’s own calculation there are a million times a trillion malaria cells needed for resistance to arise, which in scientific notation is 1018. On a log scale that’s a stone’s throw from Nicholas White’s estimate of 1020 cells per origin of resistance that I have been citing, literally an astronomically large number (there are only a paltry hundred billion, 1011, stars in our galaxy). So let me just say thank you and welcome aboard to Professor Moran. Unfortunately, he seems not to have realized the import of his calculation at the time, and has shown no enthusiasm for exploring it much after it was brought to his attention by a commenter.
Right after his calculation Moran writes "We know that the right pair of mutations … is not sufficient to confer resistance to chloroquine so the actual frequency of chloroquine resistance is far less." Far less? Far less than 1 in 1018? Now, it’s true that at least four mutations have been found in all known resistant strains of malaria. And it’s true that, although Summers et al. showed two mutations are necessary for pumping chloroquine at a low level, they might not be sufficient for chloroquine resistance in the wild. Nonetheless, a need for further mutations would only make the problem for Darwinism much worse. It wouldn’t make it better. Let me emphasize: Professor Moran’s own reasoning would make the problem much more severe than I myself have ever argued. Yet he doesn’t take any time on his blog to explore the ramifications of his own reckoning. Why doesn’t he think that’s an interesting result? Why not ponder it a bit?
Moran doesn’t seem to actually have much confidence in his own numbers. He asks the readers of his blog to help him correct his calculations — which is a commendable attitude but makes one wonder, if he’s so unsure of the likelihood of helpful combinations of mutations, whence his trust in mutation/selection? In response to the commenter who alerted him to the huge number of parasites in a million people he writes, "This is why meeting the Behe challenge is so difficult. There are too many variables and too many unknowns. You can’t calculate the probability because real evolution is much more complicated than Behe imagines." But, again, if he thinks everything is so darn complicated and incalculable, on what basis does he suppose he’s right?
That’s the reason I issued the challenge in the first place. In my experience almost all Darwinists and fellow travelers (Professor Moran doesn’t consider himself a Darwinist) simply don’t think quantitatively about what their theory asks of nature in the way of probability. When prodded to do so, they quickly encounter numbers that are, to say the least, bleak. They then seem to lose all interest in the problem and wander away. The conclusion that an unbiased observer should draw is that Darwinian claims simply don’t stand up to even the most cursory calculations.
Another commenter at Sandwalk didn’t like Moran’s calculation, so came up with his own. Great! The more the merrier! He also assumed the first mutation to be neutral, but kept a more careful accounting of its accumulation through the generations and ended up with a result of one necessary double-mutation per 420 patients. That actually strikes me as a more realistic value for a neutral mutation than Professor Moran’s. Now, at first blush 420 may seem much smaller than Moran’s number of a million patients, but that’s only because we haven’t yet considered the factor of a trillion parasites per patient. When we multiply by 1012 to get the total number of parasites per double mutation, the commenter’s odds turn out to be 1 in 1014.6 versus Moran’s 1 in 1018, again not all that far on a log scale. Either or both of these values can easily be reconciled to White’s calculation of 1 in 1020 by tweaking selection coefficients or by inferring that a further mutation is needed for effective chloroquine resistance in the wild, as Professor Moran noted.
What if the first necessary mutation isn’t neutral? What if — as seems very likely from the failure of malaria cells with one required mutation (K76T) to thrive in the lab — the first mutation is rather deleterious? The commenter estimated that, too (and also added another consideration, a selection coefficient), and came up with a value of one new double mutant per 818,500 patients. Let’s relax the admirable precision a bit and round the number up to a million. That’s the same count Professor Moran got in his (supposedly neutral) calculation, which we saw means there is one new origin per 1018 malarial parasites — not far at all on a log scale from White’s number that I cited.
The bottom line is that numbers can be tweaked and a few different scenarios can be floated, but there’s no escaping the horrendous improbability of developing chloroquine resistance in particular, or of getting two required mutations for any biological feature in general, most especially if intermediate mutations are disadvantageous. If a (selectable) step has to be skipped, the wind goes out of Darwin’s sails.
{kind=link}