 Intelligent Design
Intelligent Design
Engineering at Its Finest: Bacterial Chemotaxis and Signal Transduction
ID theorists have long urged that the case for design is both a positive and scientific argument, based on standard principles of abductive scientific reasoning. Key to the detectability of design are particular characteristics that intelligent agents often leave behind as hallmarks of their activity. We know that intelligent causes are the only category of explanation with the ability to visualize, and ultimately actualize, a complex and functionally specified end goal. Hence, presented with a complex and functionally integrated system in nature, we can infer that some measure of conscious or rational deliberation was employed in its development.
I have previously drawn attention to the engineering genius of the flagellar assembly apparatus, highlighting the clear design hallmarks pertinent to the system. For details on the design of flagellar assembly, please see my articles here and here. Now, I want to highlight yet a further clear incidence of design: the elegant molecular mechanisms underpinning flagellar rotational switching and signal transduction.
Before we can properly appreciate the details and technicalities of this system, it is necessary to take a step back and understand the foundational principles upon which it is based. Bacteria are able to move towards a food source, such as glucose, by a process known as “chemotaxis.” A requisite for this process to work is the ability of the bacterial flagellar motor to literally shift gears so that it switches from spinning counter-clockwise to rotating clockwise. This change in rotation is brought about in response to chemical stimuli from the cell’s exterior. These chemical signals are detected by a two-component signal transduction circuit that operates to induce the switch in flagellar rotation.
What is a Two-Component Regulatory System?
In general, a two-component regulatory system comprises an integral membrane protein known as a “histidine protein kinase,” and a cytoplasmic protein known as a “response regulator.” To better visualize what this looks like, take a look at the following graphic.
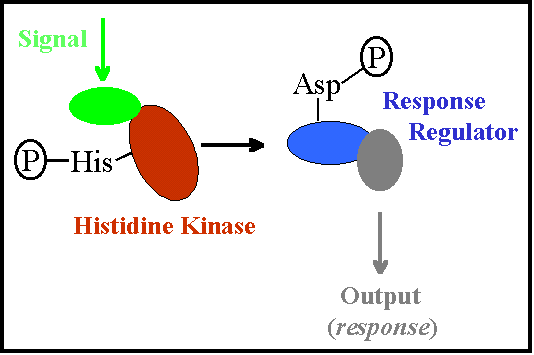
The histidine protein kinase has two domains: an input domain and a transmitter domain. The former is located on the outside of the cell, and is ideally situated to detect incoming environmental signals. The latter is situated on the cytoplasmic face of the cell membrane, and is positioned such that it can interact with the response regulator.
An external stimulus causes a conformational change in the histidine protein kinase. This causes the transfer of phosphoryl groups (autophosphorylation) from ATP to a conserved histidine residue. This phosopho-group is then moved to an aspartate residue of the response regulator. This enables the response regulator to bind to the DNA in order to regulate the transcription of its target genes.
Signal Transduction: How Bacteria Detect Food Sources
What I have thus far described represents a very basic two-component regulatory system. It was, however, necessary to look at the system in principle before we describe its application in the case of chemotaxis. It is to the latter that I now turn. Readers may find it helpful to refer to the following diagram while reading the descriptions that follow:
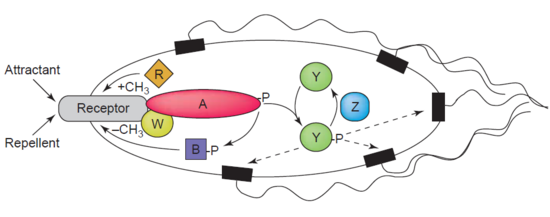
How do bacteria detect a chemical gradient? The answer lies in a certain class of transmembrane receptors called methyl-accepting chemotaxis proteins (hereafter, MCPs). Different MCPs can detect different types of molecules, and are able to bind attractants or repellents. These receptors then communicate with — and activate — the so-called “Che proteins”.
Proteins called CheA and CheW are bound to the receptor. The former is the histidine kinase for this system. Upon activation of the receptor, the CheA’s conserved histidine residue undergoes autophosphorylation. There are two response regulators called CheB and CheY. There is a transfer of a phosphoryl group to their conserved aspartate residue from CheA. CheY subsequently interacts with the flagellar switch protein called FliM. This induces the switching in flagellar direction from counter-clockwise to clockwise.
This clockwise rotation upsets the entire flagella bundle and causes it to break up. The result is that the bacterium “tumbles.” This means that bacteria are able to re-direct their course and repeatedly re-evaluate and adjust their bearings in response to environmental stimuli such as food or poisons.
As for the other response regulator I mentioned, CheB, what does it do? This is where it gets really good. When CheB is activated by the histidine kinase CheA, it operates as a methylesterase. This means that it actively removes methyl groups from glutamate residues on the receptor’s cytoplasmic surface. Meanwhile, another protein (called CheR) actively adds methyl residues to these same glutamate residues: that is to say, it works as a methyltransferase.
At this point the engineering shows a stroke of genius. If the stimulus is at a high level, there will be a corresponding decline in the level of phosphorylation of the CheA protein: and, as a consequence, of the response regulators CheY and CheB as well. Remember that the role of CheB is to remove methyl groups from glutamate residues on the receptor’s cytoplasmic surface. But now, phosphorylated CheB is not available and so this task is not performed. The degree of methylation of the MCPs will thus be raised. When the MCPs are fully methylated, the cell will swim continuously because the MCPs are no longer responsive to the stimuli.
This entails that the level of phosphorylated CheA and CheB will increase even when the level of attractant remains high, and the cell will commence the process of tumbling. But now, the phosphorylated CheB is able to demethylate the MCPs, and the receptors are again able to respond to the attracting chemical signals. In the case of repellents, the situation is similar — except that it is the least methylated MCPs which respond least while the fully methylated ones respond most. This kind of regulation also means that the bacterium has a memory system for chemical concentrations from the recent past and compares them to its currently receiving signals. It can thus detect whether it is moving towards or away from a chemical stimulus.
Conclusion
Darwinists often tell us that there is no such thing as a positive indication of intelligent design, and that our program is entirely a negative assault on the efficacy of evolution. But this argument has never been very convincing.
The system that I have described above certainly looks strikingly like a system that has been brilliantly engineered for a purpose. As rational and conscious agents ourselves, we have first-hand cause-and-effect experience of the types of systems that can be formed by virtue of blind and undirected natural processes of chance and necessity. We also have experience of systems that require the input and direction of a conscious agent. How random mutations, coupled with natural selection, could have assembled such a finely set-up system is a question to which I defy any Darwinist to give a sensible answer.