 Evolution
Evolution
 Life Sciences
Life Sciences
Understanding Ontogenetic Depth, Part I: Naming Versus Measuring
I was supposed to do this a year ago — well, long before that, too — but a glacier passed me on the interstate, and then I ran out of gas, got so depressed that I threw my notes into a box, and…oh, never mind. Let’s get started.
After the second entry in this series (part II), we’ll open up the comments section for your responses. Complete citations and additional reading will also be compiled at the end of part II.
1. Introduction: Why A Biological Distance That’s Currently Impossible to Measure, Ontogenetic Depth, Nevertheless Really Matters to Evolutionary Theory
The theory of natural selection provides a mechanistic, causal account of how living things came to look as if they had been designed for a purpose.
— Richard Dawkins (1982, 45)
Natural selection tells us absolutely nothing about underlying mechanisms of genomic changes, or their consequences on developmental changes which lead to evolutionary innovations. In short, it is silent about the emergence of complex forms.
— George L.G. Miklos (1993, 24)
How can a theory of evolution that purports to explain how creatures with trillions of cells arose from multicellular beginnings be taken seriously if all it tells us is that differential rates of destruction can alter the genetic composition of populations?
— Wallace Arthur (2004, 36)
A strong conclusion is that the evolutionary process generating the genomic programs responsible for the developmental formulation of basic eumetazoan body plans was in many ways very different from the evolutionary changes that can be observed at the species level in modern animals.
— Eric Davidson and Douglas Erwin (2010, 1)
Those statements tell you where this ontogenetic depth (OD) blog series is going. Here’s my thesis: The theory of evolution by natural selection does not explain the origin of animal form, because natural selection cannot account for origin de novo of the developmental stages required to construct (i.e., evolve) animals. The concept of OD helps us to understand why.
But first I have to eat some Corvus brachyrhynchos, partly served up 7 years ago by P.Z. Myers, but mostly stumbled on by me shortly thereafter, when I tried to make the first formulation of OD work. I am grateful to Myers for his original ammonia-capsule-under-the-nose critique, and for keeping the pressure on since then, including his annual April 7 lampoon, “Paul Nelson Day.”
As I shall explain in part II, however, I don’t think Myers grasps the severity of the problem posed by ontogenetic depth. As evidence, I’ll describe a long conversation he and I had at the Society for Developmental Biology 2004 meeting, in Calgary, at a session where I was presenting a poster. I’m quite sure Myers remembers the conversation and a little picture he drew for me on a piece of scrap paper.
But that’s for later. To start, I want to show why OD is easy to understand — in one sense, anyway — but, at the moment, nigh-well impossible to measure. Mmm, yummy crow feathers. The punch line after the feathers, however, may be counterintuitive to readers, but here it is anyway: not being able to give an exact value to a distance doesn’t mean one can ignore it in constructing theories or explaining transitions. For the macroevolution of animals de novo to occur via natural selection, ontogenetic depth must be traversed. This is an inescapable, and currently unsolved, problem. And every improvement in measuring OD will only make the problem more challenging.
2. A Useful Distinction: Naming versus Measuring
Right now, the skies are clear in Chicago. Looking into the southeastern sky, I can see the sun, some distance away in space.
Now, let’s suppose that we’re looking at the sun 500 years ago, before Kepler, Newton, Huygens, and Cassini. We know that the sun is some measurable distance from the Earth, and we can give that distance a name. Call it the Earth-to-Sun distance, or, as it now known, the Astronomical Unit (AU; see Figure 1).

Figure 1
So how far away, exactly, is the Sun from the Earth? Five hundred years ago, we didn’t have telescopes, radar, satellites, or any of the other instruments currently used to measure the AU. But we did have geometry, and could grasp the concept of parallax. It should be fairly easy, using naked-eye observations of the Sun and the planets, to estimate the exact value of the AU.
Not really. As the history of astronomy shows, measurements (or estimates) of the AU, attempted since antiquity, didn’t settle on anything close to the modern value until the late 18th century. The data required to perform the triangulation depicted in Figure 2, accurately, by employing the Transit of Venus, were simply not available, or were known too imprecisely to be useful.
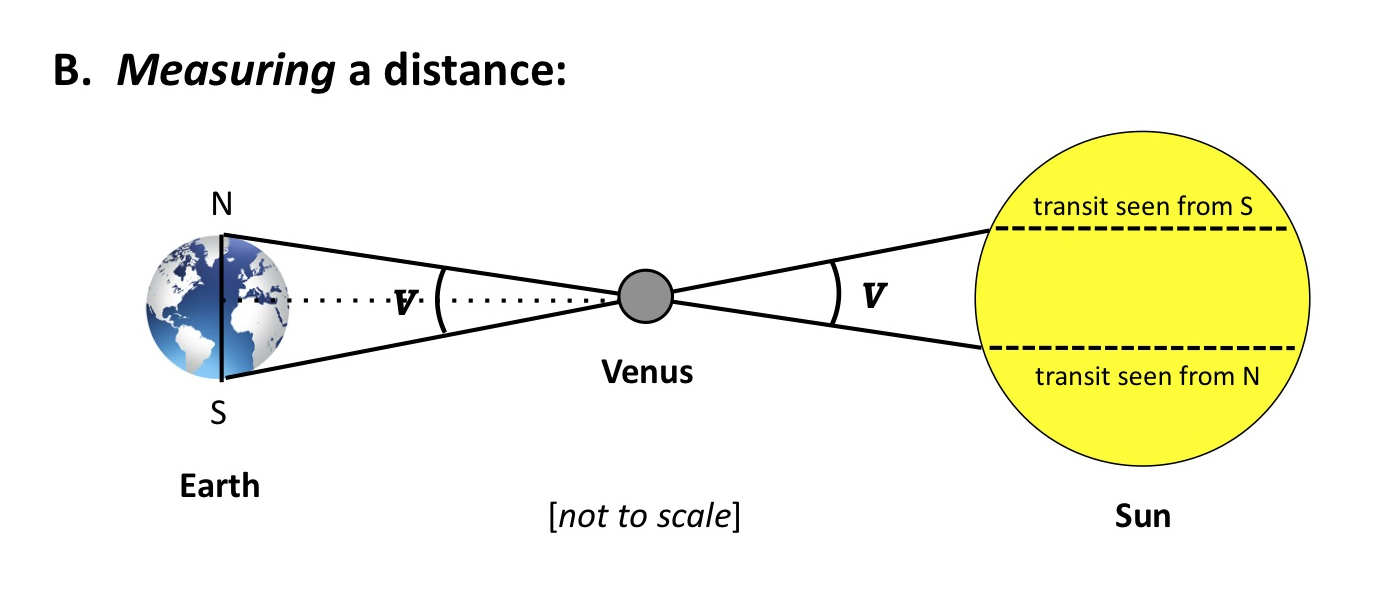
Figure 2
Does that mean the concept of the AU is unintelligible? Of course not. The difficulties we face in measuring a distance do not entail that the distance itself is unreal or unimportant. The difficulties just mean that the distance may be tricky to specify in exact terms.
3. Ontogenetic Depth: Naming versus Measuring
Segue to ontogenetic depth. Here is my original formulation (2003, 459):
Ontogenetic depth estimates the distance, in terms of cell division and differentiation, between a unicellular condition and the macroscopic adult metazoan capable of reproduction.
The words in bold define, or name, a distance. The words in italics, by contrast, represent an attempt (and a poor one at that; thanks, Prof. Myers, for showing as much) to measure that distance. If the reader can grasp this distinction, namely, between defining or naming a distance, versus measuring it, much of the confusion surrounding ontogenetic depth dissolves away. The important point to remember is this: the distance (OD) is real, and must be traversed by any candidate evolutionary process, whether we can measure OD precisely or not.
Consider a parallel to defining versus measuring the AU. Figure 3 shows OD as the distance between a fertilized C. elegans egg, and the reproductively capable adult phenotype. We can think of this like looking at the Sun, and knowing it is some distance [finite interval] away in space.
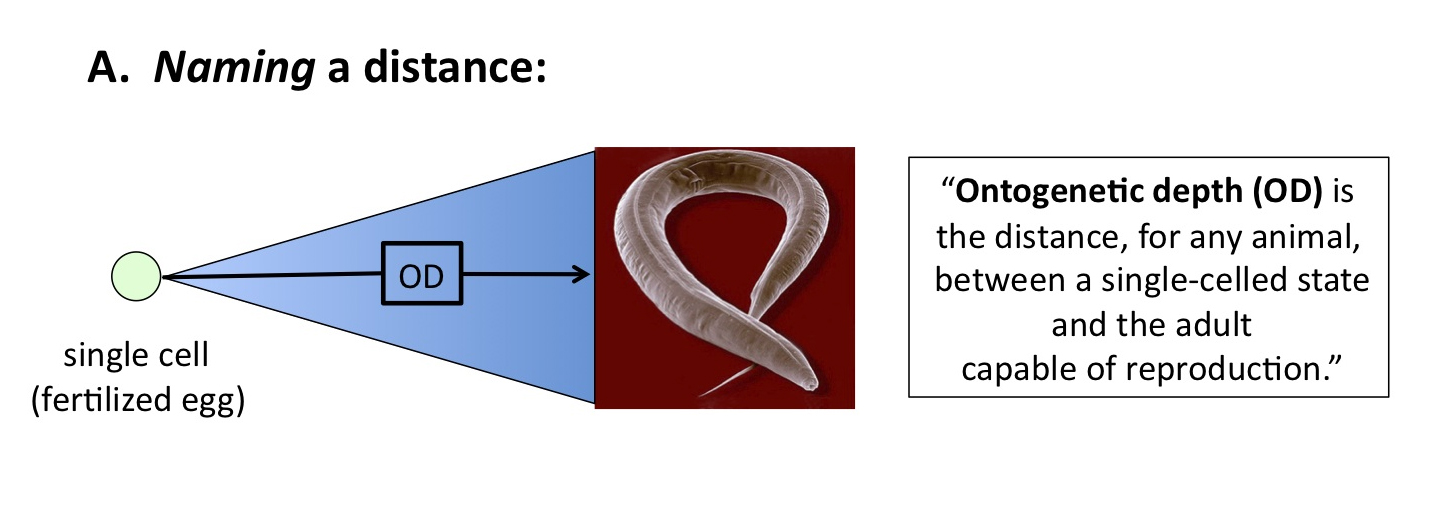
Figure 3
Figure 4, by contrast, posits that cell division and differentiation (i.e., cell types) provide the correct units for measuring — i.e., giving an exact value — to OD:
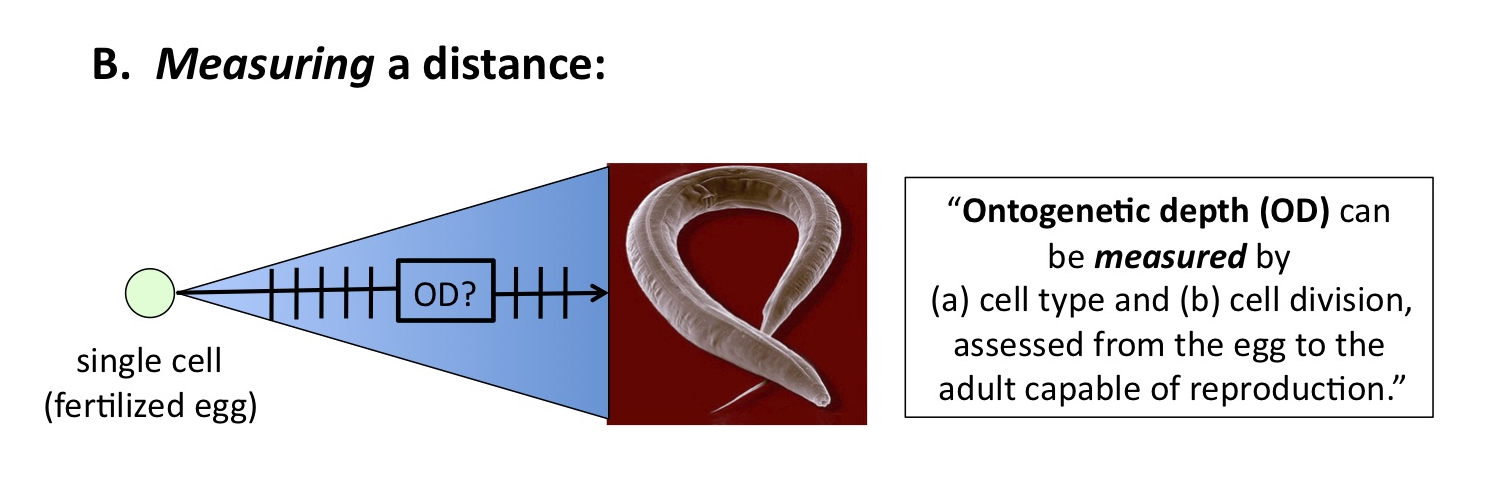
Figure 4
See the difference? Anyone would agree that there’s a distance between egg and adult in C. elegans. The question is how best to measure that distance, in units (or metrics) that allow comparison with other animal groups. In his original critique, Myers gave some reasons for thinking that the metrics of cell type and cell division wouldn’t work. But he undersold the difficulty, as I’ll illustrate next.
Here’s the cell lineage (see Figure 5) of Toy Organism Alpha, with 16 cells of 5 types. Four rounds of cell division separate any cell in the adult from the starting point, the fertilized egg. A simple metric of OD for this toy organism would multiply the average number of cell divisions, between the egg and any cell in the adult (4), times the total number of cell types (5, with the red cell as the germ line producing gametes), to yield an OD of 20.
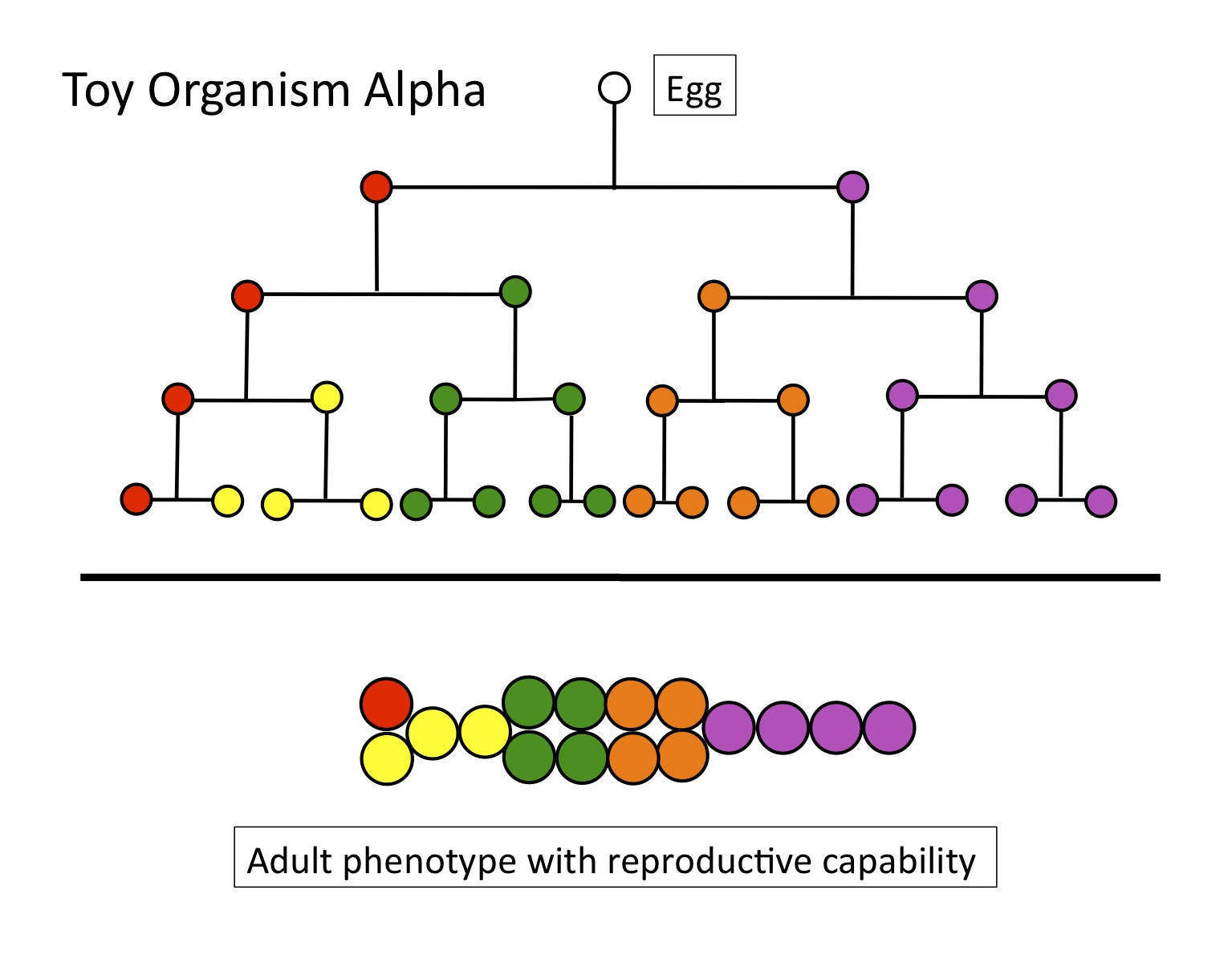
Figure 5
But compare Toy Organism Alpha to Toy Organism Beta. While they have the same OD — if we use the simple metric of 4 rounds of cell division X 5 cell types = 20 — their respective adult forms are very different. Beta has a large internal cavity, and places its “gonad” (the red cell) within the wall of that cavity, whereas Alpha’s gonad is exterior. The positions of other cells differ as well.
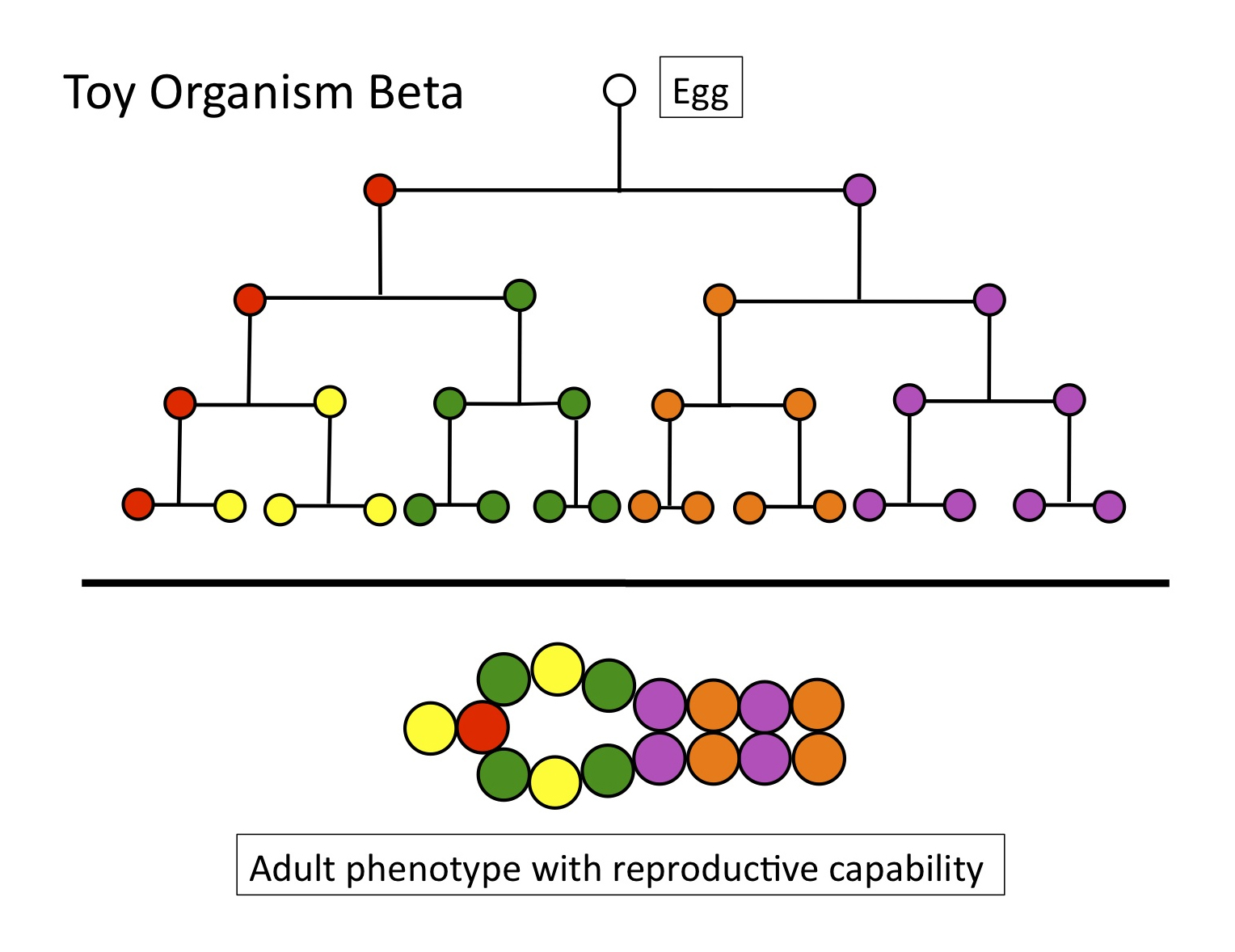
Figure 6
If reproductive capability depends (at least in part) on precise form, as it does for any real animal, then measuring ontogenetic depth will require specifying the three-dimensional positions and relationships of individual cells or tissues. This positional information cannot be captured either by cell division (lineages) or cell type, and providing a metric for it will be enormously more difficult.
“So why bother about a distance that cannot yet be measured?” the reader is probably asking. “What’s the point?”
4. The Theory of Evolution by Natural Selection Itself Requires That We Explain Ontogenetic Depth
That’s why. As I’ll explain in Part II, using the process of natural selection in evolutionary explanation places strict evidential demands on the investigator. Evolving the goal-directed structures of metazoan development de novo raises challenges that natural selection — because the process lacks foresight — may in principle be incapable of explaining.