 Evolution
Evolution
 Intelligent Design
Intelligent Design
Confusing Evidence for Common Ancestry With Evidence for Darwinian Evolution
 Download the Complete “Truth or Dare” with Dr. Ken Miller Lecture Guide Permission Granted to Copy and Distribute for Educational Use. Links to our 7-Part Series Responding to Ken Miller: • Part 1: Science and Religion: Is Evolution “Random and Undirected”? |
Update 8/7/13: Since this response to Ken Miller was posted, even stronger evidence of function for the beta-globin pseudogene has been reported in the scientific literature, further refuting Miller’s argument. See Dover Revisited: With Beta-Globin Pseudogene Now Found to Be Functional, an Icon of the “Junk DNA” Argument Bites the Dust.
Behe puts it even more clearly in Darwin’s Black Box: “Although useful for determining lines of descent…comparing sequences cannot show how a complex biochemical system achieved its function–the question that most concerns us in this book. By way of analogy, the instruction manuals for two different models of computer put out by the same company might have many identical words, sentences, and even paragraphs, suggesting a common ancestry (perhaps the same author wrote both manuals), but comparing the sequences of letters in the instruction manuals will never tell us if a computer can be produced step-by-step starting from a typewriter….Like the sequence analysts, I believe the evidence strongly supports common descent. But the root question remains unanswered: What has caused complex systems to form?”18Both at the Dover trial and in his lectures and books (such as Only a Theory), one of Dr. Kenneth Miller’s primary responses to Michael Behe’s arguments for irreducible complexity is to cite evidence for common ancestry. This class of evidence does not refute Behe because at most, evidence of sequence similarity in DNA demonstrates common ancestry–not a Darwinian evolutionary pathway. Indeed, on closer inspection, it turns out that much of Miller’s favorite evidence does not even provide a strong case for common descent: Miller assumes that functional genetic similarities must result from common descent, ignoring the possibility that such biochemical similarities might result from common design upon a functional blueprint.
First, one of Miller’s most common mistakes is to forget that evidence of common ancestry is NOT evidence of a Darwinian pathway, and thereby does not refute irreducible complexity. Behe, the leading proponent of irreducible complexity who also accepts common descent, aptly observes that “modern Darwinists point to evidence of common descent and erroneously assume it to be evidence of the power of random mutation.”17
Miller’s citation of similarities in DNA sequences in no way refutes irreducible complexity, nor does it demonstrate a stepwise Darwinian evolutionary pathway.
| C. Truth or Dare: Why does Dr. Miller repeatedly offer evidence of common descent as if it refutes irreducible complexity, when it doesn’t logically demonstrate a Darwinian pathway and in fact the leading proponent of irreducible complexity accepts common descent? |
Second, even though intelligent design is not necessarily incompatible with common descent (more on this later in Section IV), it should be noted that many of Dr. Miller’s centerpiece examples of evidence for common descent turn out to be quite weak.
As noted, functional genetic similarities may result from common design rather than common descent. After all, designers regularly re-use components or parts that work in different designs–such as re-using cars and wheels in airplanes, or re-using keyboards on both laptops and cell phones. Thus, when we find functional genetic similarity in different organisms, it might indicate common design.
Though he might not admit it, some of Miller’s arguments implicitly concede this point. Miller contends that the way to refute design is not to find shared functional similarities but to find supposed nonfunctional “junk” DNA. As Miller writes: “Intelligent design cannot explain the presence of a nonfunctional pseudogene, unless it is willing to allow that the designer made serious errors, wasting millions of bases of DNA on a blueprint full of junk and scribbles. Evolution, however, can explain them easily. Pseudogenes are nothing more than chance experiments in gene duplication that have failed, and they persist in the genome as evolutionary remnants of the past history of the b-globin genes.”19
Though Miller wrote those words in 1994, he continues to use the beta-globin pseudogene as a refutation of ID–it was his centerpiece example of a pseudogene in his 2005 Dover testimony, in his 2008 book Only a Theory, and it’s often mentioned in his lectures. Privately, Miller has cited such pseudogenes as “case-closed” evidence of common descent because “common ancestry is the only possible explanation for so many matching errors in the same gene.”20
Dr. Miller may be closing this case prematurely. Two authors wrote in Annual Review of Genetics: “pseudogenes that have been suitably investigated often exhibit functional roles.”21 According to these authors, functions include “gene expression, gene regulation, [and] generation of genetic (antibody, antigenic, and other) diversity.”21 They further suggest that conserved DNA sequences in pseudogenes implies they have function: “Pseudogenes exhibit evolutionary conservation of gene sequence, reduced nucleotide variability, excess synonymous over nonsynonymous nucleotide polymorphism, and other features that are expected in genes or DNA sequences that have functional roles.”21 Following such sound logic, the British pro-ID group Truth in Science recounts how Miller’s favorite example — the beta-globin pseudogene — shows evidence of conserved sequence, implying that it could have function, which would refute Miller’s centerpiece evidence of a functionless, junk DNA “pseudogene”:
“The very fact that the beta-globin pseudogene appears to be conserved in humans, chimpanzees and gorillas speaks eloquently of the fact that this DNA has some important biological function. Genetic sequences are conserved and maintained when any mutation would render them non-functional (or less functional) and when any loss of activity is damaging the organism’s prospects of survival. Such sequences are said to be under purifying (or stabilising) selection which means that deleterious mutations are removed from the gene pool restricting genetic diversity. … According to the recent review by Sasidharan and Gerstein: ‘Although pseudogenes have generally been considered as evolutionary ‘dead-ends’, a large proportion of these sequences seem to be under some form of purifying selection – whereby natural selection eliminates deleterious mutations from the population – and genetic elements under selection have some use.’ In the case of the beta-globin pseudogene, Wanapirak et al. have reported amazing conservation in the fine structure of the DNA with identical super-helical twists in the human, mouse, bovine, rabbit and chicken genomes. It needs to be remembered that maintenance of the genetic integrity of these structures is biochemically costly. It takes energy to duplicate DNA. The replicating machinery in the cell has built-in proof reading and excising enzymes that constantly check for mutation and damage. Numerous repair mechanisms have been identified to correct genetic damage and to excise incorrect sequences.”22
By assuming that the pseudogenes like the beta-globin pseudogene in humans are functionless “junk” DNA, Dr. Miller is not only wrong; he may be hindering the progress of science by discouraging scientists from discovering its true function. This is ironic for someone who has accused ID of stopping science.
Finally, a piece of evidence Dr. Miller commonly cites as demonstrating human/chimp common ancestry is the fusion of chromosome 2 in humans, which he argues has a structure similar to what one would expect if chimp chromosomes 2a and 2b were fused together, end to end. Without belaboring the details (which are covered elsewhere23), the evidence for human chromosomal fusion simply indicates that our ancestors once had 48 chromosomes. But it tells us nothing definitive about whether our lineage leads back to a common ancestor shared with with apes. Human chromosomal fusion merely shows that at some point within our human lineage, two chromosomes became fused. That’s it.
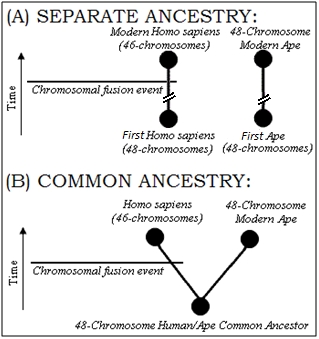
If we step outside the Darwinian box, then the following scenario becomes possible: (1) The human lineage arose separately from that of apes with 48 chromosmes, (2) a chromosomal-fusion event occurred, and (3) the trait spread throughout the human population. In such a scenario, the evidence would appear precisely as we find it, without any common ancestry between humans and apes. The two diagrams at right show two models for explaining the evidence for human chromosomal fusion.
At most, the fusion evidence confirms something we already knew: humans and apes share a similar genetic structure. But this might have been predicted by morphological studies without considering evolution. Again, common design can also account for such functional genetic similarities, and the fusion evidence does not demonstrate that humans share a common ancestor with apes.
Dr. Miller may reply that his model predicts the fusion evidence. But if we didn’t find evidence for fusion in human chromosome 2, would that really refute Darwinism? No. Evolutionists would just claim that the fused telomeres and extra centromere were deleted.
| D. Truth or Dare: Has it actually been established that pseudogenes–especially those with conserved sequence like the beta-globin pseudogene–are functionless “junk”-DNA? Wouldn’t it be more appropriate to take a “wait and see” approach, especially since so many types of once-dismissed “junk”-DNA have turned out to have function? Why must common design be excluded from our explanatory toolkit to account for the genetic similarities between humans and apes? Does the fusion evidence really require we share a common ancestor with apes? |
[Editor’s Note: Ken Miller speaks regularly on intelligent design (ID), and for years has repeatedly promoted the same misrepresentations of ID when speaking on the topic. This is Part 3 of a series of posts that comprise a lecture guide for those listening to lectures by Dr. Miller against ID. When this series is complete, the entire lecture guide will be released as a single document. Also, don’t forget to read Dover Revisited: With Beta-Globin Pseudogene Now Found to Be Functional, an Icon of the “Junk DNA” Argument Bites the Dust, which is a more recent discussion of the strong evidence of function now known for the beta-globin pseudogene.]
References Cited:
[17.] The Edge of Evolution, p. 95 (2007).
[18.] Darwin’s Black Box, pp. 175-176 (1996),
[19.] “Life’s Grand Design,” Technology Review, Vol 97(2): 24-32 (February / March 1994).
[20.] Private correspondence with Dr. Miller.
[21.] Evgeniy S. Balakirev, and Francisco J. Ayala, Pseudogenes, “Are They “Junk” or Functional DNA?,” Annual Review of Genetics, Vol. 37:123–51 (2003), emphasis added.
[22.] “The Changing Face of Pseudogenes,” Truth in Science (internal citations removed).
[23.] See And the Miller Told His Tale: Ken Miller’s Cold (Chromosomal) Fusion (Updated)
or Weird Science